|
MENU

|
| FNCA 2002 WORKSHOP ON MUTATION BREEDING |
| The Progress and Potentials
of Mutation Induction in Vegetatively Propagated Plants
in Malaysia.
Mohd. Nazir Basiran and Sakinah Ariffin
Malaysian Institute for Nuclear Technology Research (MINT)
|
| |
|
|
|
|
Plants can be vegetatively propagated either in
vivo or in vitro. These vegetative propagation techniques are commonly
applied for cloning of plants that have highly segregating seeds,
long juvenility or are parthenocarpic. Plants such as pineapples
and bananas for example, are vegetatively propagated because it
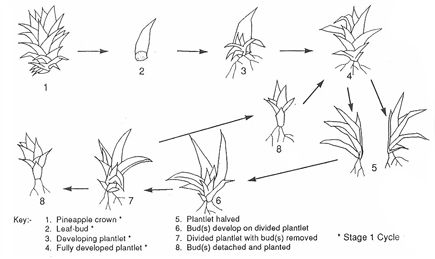
is not preferable to have seeded fruits. In pineapple, leaf budding
has been shown to be a good way of mass propagating in vivo (Fig.
1). An average of 15 leaf-buds can be obtained from a single crown.
Meanwhile, suckers have traditionally been used to propagate banana.
However, the rate of in vivo propagation through suckers or leaf
budding is not sufficiently rapid and does not have growth uniformity
for commercial scale production. Therefore, micropropagation through
tissue culture techniques is developed and has been very instrumental
in improving the rate of clonal propagation as well as the quality
of both pineapple and banana seedlings.
For fruit trees such as durian (Durio zibethinus), mango, rambutan,
ciku (Achras sapota L.) and citrus, budding and marcotting are the
traditional methods for propagation. Other methods such as stem
cutting and cleft grafting have been successfully developed for
duku langsat (Lansium domesticum), durian and ciku. These are important
development not only for clonal propagation of new varieties but
also useful for a genetic improvement program of these fruit trees.
An efficient vegetative propagation system whether in vivo or in
vitro is very pertinent prerequisite for genetic improvement procedures
of such plants which have long juvenility, difficult sexual compatibility,
and sterility. Physical and chemical mutagen treatments can induce
mutations on the growing points of these vegetatively propagated
plants. The mutated sectors on these treated tissues can then be
grown and isolated from the non-mutated sectors through a series
of vegetative propagation procedures either in vitro or in vivo.
Many useful mutant cultivars with improved agronomic characteristics
have been developed through this methodology.
The advent of tissue culture technology enables small and easily
handled amount of tissue to be treated. Excised stem tips or callus
growing on standard nutrient medium in a test tube for examples,
could be treated and the explants grown to maturity and evaluated
for useful mutations. Advanced in vitro manipulations such as somatic
embryogenesis and single cell cultures also reduce the problem of
chimerism often encountered in the induction of mutations on vegetatively
propagated plants. |
|
|
|
|
Fig. 1: Illustration of the leaf-budding
cycle from Wong, W.W.W (1992). |
|
|
The varietal improvement activities in Malaysia have adopted both approaches of in vivo and in vitro vegetative propagation for mutagenesis in plants such as banana, orchids and other foliage ornamentals. The objectives are to develop varieties that have better agronomic characteristics such as high yield, disease resistance or tolerance, better and attractive flower colour, longer shelf life and good plant architecture. |
|
| |
|
|
|
| Pisang Berangan |
|
| The interests in banana mutagenesis was created when location trials of gamma irradiated GN-60A seedlings produced the early maturing, high yield and better agronomic characteristics mutant clone called NOVARIA. Subsequently, a multi-institutional and multi-disciplinary research group was formed to tackle issues of disease particularly Fusarium wilt, to improve a local cultivar P. Berangan through in vitro mutagenesis. The agronomic characters considered as target include short plant height, earliness in fruiting and high yielding capacity. In general, gamma induced mutations caused morphological changes and increase in variability of quantitative traits. A much higher frequency for two important qualitative traits was observed in early fruiting and dwarfness or short stature. Other variants which showed an increase of up to 6 folds include leaf characteristics such as uneven lamina, size and shape, changes in leaf coloration and characteristics of the pseudostem. |
|
| Pisang Mas |
|
| Another banana of interest is Pisang Mas. Pisang Mas which is a desert banana belonging to the AA group has great export potentials. However, the fruits are small with thin skin and have low bunch weight. A breeding strategy to improve these characteristics was formulated to produce tetraploid which can then be used for crossings with diploid to produce secondary triploid. The polyploidisation was achieved by using colchicines treatment on meristem pieces explanted from in vitro regenerated shoots. Colchicine treatments in the range of 0.25 to 2.0% at 1 to 4 hrs produced 7.4% tetraploids 0.42% octoploid and 29.65% mixoploid. However, there was no significant trend of polyploidisation corresponding to the duration or concentration of treatments. In general, higher concentration of colchicines produces higher rate of polyploidisation. The highest frequency of tetraploid induction was at 1.0% colchicines treatment for 3 hrs. |
|
| The ploidy level was determined by flowcytometry (Fig. 2) and also by observations on the morphological characteristics. Generally, the polyploid seedlings have bigger pseudostem, shorter stature with thicker and smaller leaves and slightly darker green. The measurement of the stomata size and density were also taken from imprint of leaves of 2 month old in vitro plantlets. These are good and rapid estimation of ploidy level. |
|
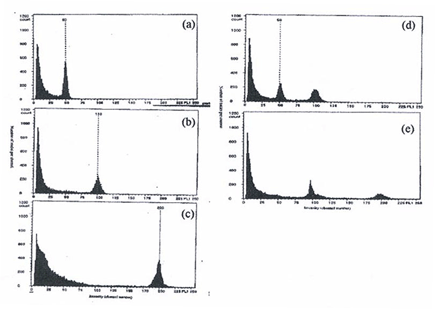 |
| Fig. 2: Histogram of treated Pisang Mas (a) Control; (b) tetraploid; (c) octoploid;
(d) mixoploid 2n + 4n; (e) mixoploid 4n + 8n. |
|
| Somaclonal variations |
|
| Besides the induced mutation activities, new variants were also obtained through somaclonal variations. The occurrence of somaclonal variations amongst in vitro propagated seedlings can be as high as 60% depending on the cultivars. Although most of the variations are undesirable agronomically, the phenomenon can
be a source of genetic variation. Through careful selection, variants with earliness in fruiting, fruit color (P. Udang) and Fusarium wilt tolerance (P. Rastali) have been obtained. Selection on Fusarium ‘hot spot’ and comparison between Goldfinger, P. Berangan, and NOVARIA have shown that one P. Rastali somaclonal variant perform as good as the Goldfinger and has high tolerance towards Fusarium. Further propagation of the variant has produced highly tolerant plants. |
|
| |
|
|
| |
| Ornamental foliage is one group of plants that benefit tremendously with the advancement of vegetative propagation technology. The technology helps to expedite development of new plants when combined with the induced mutation technology. The method accelerates the chance of natural mutation to occur by producing leaf chimeras, ‘sports’ and other mutations. Although many of the mutants would be similar to variants already in existence, possibilities exist for the development of a new variant. The same technology is also applicable to flowering ornamental plants to change flower colour and shapes. |
|
| Orchids |
|
| Orchids as a major cut-flower export commodity of Malaysia have not had any significant input of new and attractive varieties that can elevate the export values to a new plateau. Although this may not be the only factor in impeding a greater expansion of the floriculture industry, the availability of wide range of attractive varieties will give a competitive advantage for growers. Induced mutation techniques have been seen as an alternative or complementary tool for the development of new orchid varieties. |
|
| Results of exploratory experiments to assess the feasibility of using ionizing radiation to induce mutations in orchids found that different genus and species responded differently to the irradiation treatments. The radiosensitivity seemed to be influenced by the age of the protocorms and differences in ploidy level of different species or hybrids. Generally, irradiation treatments affect the characteristics of the plant and flowers as well as the growth. Higher doses stunted growth of the plant and caused severe morphological changes. High frequency of protocorms irradiated at high doses developed very small stunted shoots with large stumpy roots. |
|
| Numerous combinations of colour and shape variations in the petals, sepals and lip of the flower were observed. One mutant has totally white flowers but with similar flower shape as the control, indicating single characteristic change. The different color variations are expressed as whole or in gradual distribution of intensity from pale to darker shade in the petals or sepals. The intensity and uniformity of the pigment distribution pattern may also vary in a single flower resulting in colour patches. Flower size of mutant can be 20% larger or up to 30% smaller than the control. A mutant with slight aroma was also obtained. |
|
| The flower arrangement of the spray also showed some variations. The flowers are arranged alternately on each side of the stalk and can be either laterally spread almost at right angle to the stalk or at a more acute angle. Some are arranged in a more compact and tight arrangement along the flower stalk. One mutant was found to have flowers arranged closer together towards the tip of the stalk. |
|
| Irradiation treatment also affected the shelf life of the flowers. The shelf life can be extended up to 30 days as compared to the control or can be reduced by half to 8 days. Other characteristic changes or abnormalities that had been observed included variegated leaves, branching sprays, zigzag flower stalk and deformed flowers. Some of these observed abnormalities are not expressed in the subsequent flowering, indicating that they are not stable characteristic change. |
|
| Irradiation has been shown to either increase or decrease pigmentation intensity of flowers in other species as well. These changes in pigmentation intensity and distribution patterns, and the changes in the petal shapes of orchids were stably expressed in the second and subsequent flowering. The same flower characteristics were also observed in the flowers of the vegetative regenerants from the mutant plant. These changes in characteristics are due to genetic changes induced by gamma irradiation, as shown by the DNA polymorphism between selected mutants and the control as well as between the mutants. |
|
| Alpinia purpurata |
|
| Alpinias or gingers make very impressive container plants or can be planted in the garden as landscaping. They belong to the Zingiberaceae and have sturdy leaf stalks rising from the tubers underground. They can grow up to nine feet tall and produce flower which consists of layers of cerise bracts and is used in the cut flower industry around the world for floral arrangements. They are sold in 3.5 in pots rooted or rhizome depending on availability. Several hybrids with varying degree of pinkness are available with height of up to 12 feet tall. While the gingerly scented attractive flowers and aromatic leaves are unique and exotic, reduction in height of the plant is desirable to make the plant more suitable for landscaping or as potted plant. The flowers can last up to three weeks on the plant, but they only have 7-10 days of vase life. Mutagenesis is a tool that may be employed to improve architecture of the plant and the shelf life of the flowers. |
|
| This exotic tropical plant is easy to grow and requires well draining soil kept on the moist side and prefers humidity and partial sun/shade. Depending on the hybrids, they can be propagated vegetatively quite easily using rhizome segments containing three to four buds. However, fruits with viable seeds are seldom produced and large scale multiplication by cuttings is very slow. Thus, there is a need for in vitro micropropagation. Several reports of in vitro cultures of Zingiber officinale and Alpinia purpurata have been published. An in vitro propagation system was thus developed using the floral buds. |
|
| Using the in vitro propagation system the optimum dose for mutagenesis was estimated at between 5-10Gy (Fig. 3). In vitro mutagenesis of the red bract alpinia has produced mutant with pink bracts, several mutants with variegated leaves and a bushy plant. The bushy mutant plant is also dwarf and has not flowered after several years of planting in the pot. Mutants with two types of leave variegations had also been obtained. One type has leaves of green and yellow, while the other has green and white leaves. Both are very difficult to convert from tissue culture to the greenhouse. Although the green and white leaves were successfully transferred to the greenhouse, further propagation and growth maintenance were very difficult. They are mostly chimeric and the young shoots arising from the rhizome showed segregation of characteristics. |
|
| New experiments are being conducted to induce mutations by fractionated irradiation treatments of in vitro explants. Regenerated shoots were irradiated at 5Gy for three times pre-subcultures. These irradiated shoots were then allowed to divide further and transferred to the greenhouse. A high frequency of leave variegation and leave morphological variations is observed. |
|
| Foliage ornamentals |
|
| Several woody medium size shrubs which are easily propagated by nodal cuttings were subjected to mutagenesis by gamma rays. Species such as Duranta, Cordylines, Bougainvillea and Dracaena marginata were found to be able to withstand slightly higher irradiation doses than species with softer stem such as Dracaena sanderiana and Rheodiscolor. Radiosensitivity test on Cordylines cuttings has shown that the LD50 is 30Gy while for Dracaena and Rheodiscolor it is approximately 15Gy. However, lower doses are often preferred if ‘green’ or shoot cuttings of the woody species are used. Following the irradiation treatment, mutations can be observed as early as the first vegetative generation (the regenerated shoots from the irradiated axillary buds). However, these are often chimeric and cutting back is necessary to isolate the mutated sector. Mutants with leave variegations and stunted growth (dwarfs) were successfully selected from mutagenesis of Cordylines, Duranta species and Rheodiscolor. Two mutants with flower color change were obtained in Hibiscus rosa-sinensis after three vegetative generations and the mutant remained stable throughout successive generations. |
|
|
| |
|
|
|
| Induced mutation either by physical or chemical mutagens is a powerful tool for varietal development of vegetatively propagated plants. Several mutants with useful characteristics have been successfully developed in bananas and ornamental plants. The wide range of variations in flower pigmentation intensity, size and shape, and the possibilities of inducing change in only one character make irradiation-induced mutation a potential tool for further improvement of orchid and other ornamental varieties. These successes can be extended to other fruit trees especially those which can be efficiently propagated vegetatively. The induced mutation approach is particularly important for fruit trees such as mangosteen and lansium which are believed to be apomictic. |
|
|
|
|
|
| Azhar, M., Mak, C. and Mohd Nazir B. (2002). Polyploidy induction in Pisang Mas, a diploid banana (Musa spp.). MINT/L/2002/07. |
|
| Chang, B.K.W. and Criley, R.A. (1993). Clonal propagation of pink ginger in vitro. HortScience 28(12):1203 |
|
| Illg, R.D. and Faria, R.T. (1995). Micropropagation of Alpinia purpurata from inflorescence buds. Plant Cell, Tissue and Organ Culture 40:183-185. |
|
| Ling, C. H. (1998). Effects of gamma irradiation on in vitro cultures of selected orchid hybrids, MS Thesis, Universiti Putra Malaysia, Serdang, Malaysia. 113 Pages. |
|
| Mak C. and Ho Y.W., (1998). Banana improvement: somaclonal variation and in vitro mutation breeding. In Proceedings of the Third National Congress on Genetics, 18-19 Novemer 1998, UKM, Bangi. pp277-285. Eds Mohamad O. et al. |
|
| Mohd. Nazir, B., Sakinah, A., Zurinah, A. S. and Zarina, A. R. (1998). DNA polymorphisms in orchid mutants. In Proceedings of FAO/IAEA Seminar on mutation techniques and molecular genetics for tropical and subtropical plant improvement in Asia and the Pacific region. IAEA-SR-210. |
|
| Mohd. Nazir, B. (2000). In Proceedings of MINT R&D Seminar 2000. Malaysian Institute for Nuclear Technology Research, Malaysia. |
|
| Mohd. Nazir, B., and Sakinah, A. (2001). Molecular techniques as complementary tools in orchid mutagenesis. Proceedings of The 2001FNCA Workshop on Agriculture: Plant Mutation Breeding: & Biofertilizer, Aug 20-24, Bangkok, Thailand, Japan Atomic Energy Research Institute. |
|
| Shuhaimi, (1996). Ujian radiosensitiviti ke atas tanaman hiasan yang dibiak secara tampang. Cordyline fruticosa ‘Compacta’. MINT/T/1996(172). |
|
| S.T. Chong, (1992). Propagation of durian (Durio zibethinus) by cleft grafting. In Proceedings of the National IRPA Seminar (Agriculture Sector) Vol. 1 Crops and Plants. pp535-536. Eds Y.W.Ho et al. |
|
| S.T. Chong, (1992). Recent developments in vegetative propagation of some tropical fruit trees. In Proceedings of the National IRPA Seminar (Agriculture Sector) Vol. 1 Crops and Plants. pp525-526. Eds Y.W.Ho et al. |
|
| T.B. Chai, (1992). Development of vegetative propagation techniques for duku langsat.. In Proceedings of the National IRPA Seminar (Agriculture Sector) Vol. 1 Crops and Plants. pp549-550. Eds Y.W.Ho et al. |
|
| William W. W. Wong, (1992). Propagation of fruits, spices and nuts. In Proceedings of the National IRPA Seminar (Agriculture Sector) Vol. 1 Crops and Plants. pp527-528. Eds Y.W.Ho et al. |
|
|
page top↑ |